Teil 9
Heute werden wir ohne besondere Einleitung die Meiose zu Ende führen. Später fassen wir dann all das zusammen, was an dieser besonderen Zellteilung für unser Vorhaben wichtig ist. In der letzten Abbildung aus dem 3. Teil (Abb. 5) war die Anaphase-I zu sehen. In der Telophase-I wandern die beiden Chromosomengruppen zu den beiden Zellpolen und verharren dort kurz in der Interkinese, einer speziellen Interphase. Dabei werden die Chromosomen jedoch nicht entspiralisiert, sondern verbleiben in der Transportform, denn es folgt nun unmittelbar die Meiose-II. Ohne daß eine Zellwand eingebaut oder Kernmembranen gebildet werden, verhalten sich die beiden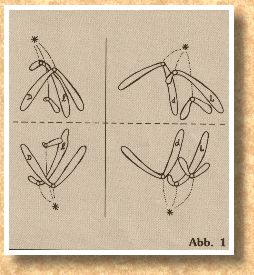 Chromosomengruppen wie zwei getrennte Zellen, die nun synchron eine mitoseartige Teilung vollziehen. Nach einer kurzen Prophase-II ordnen sich die beiden Chromosomengruppen in der Metaphase-II in zwei getrennte Aquatorialplatten an. Die beiden Teilungsebenen stehen senkrecht zu der ersten Teilungsebene, wie wir aus Abb. l entnehmen können.
Chromosomengruppen wie zwei getrennte Zellen, die nun synchron eine mitoseartige Teilung vollziehen. Nach einer kurzen Prophase-II ordnen sich die beiden Chromosomengruppen in der Metaphase-II in zwei getrennte Aquatorialplatten an. Die beiden Teilungsebenen stehen senkrecht zu der ersten Teilungsebene, wie wir aus Abb. l entnehmen können.Aus den Gameten werden Spermien
Wir sehen hier die Anaphase-II, und es sieht so aus, als ob zwei mitotische Anaphasen nebeneinander gezeichnet wären. Die beiden identischen Chromatiden eines jeden Chromosoms sind voneinander getrennt worden. Der kleine Unterschied besteht darin, daß in jeder Chromosomengruppe nur noch eines der beiden Homoigen vorhanden ist. Es geht nun ganz normal weiter. In der Telophase-II wandern die Chromosomen endgültig zu den vier Zellpolen und beginnen sich zu entspiralisieren. Es werden senkrecht zueinander zwei Zellwände eingezogen und um jede der vier Chromosomengruppen eine Kernmembran gebildet. Das Endprodukt sind vier voneinander unabhängige Zellen in der Interphase, die vier Gameten, die sich in unserem Fall getrennt voneinander zu Spermien entwickeln, denn wir haben die Meiose ja an einem Modell-Kater betrachtet. Hätten wir unsere Katze aus dem 2. Teil als Modell genommen, dann wäre aus einem der vier Teilungsprodukte eine Eizelle oder Oocyte entstanden, die übrigen drei wären zu den sog. Polkörpern verkümmert. Welches von den vier Teilungsprodukten nun zur Eizelle wird, bleibt allerdings dem schon so oft erwähnten ZUFALL überlassen. Bevor wir uns aber weiter mit dem ZUFALL befassen, wollen wir die für unsere genetischen Analysen und Berechnungen wichtigen Schritte der Meiose noch einmal wiederholen und zusammenfassen.
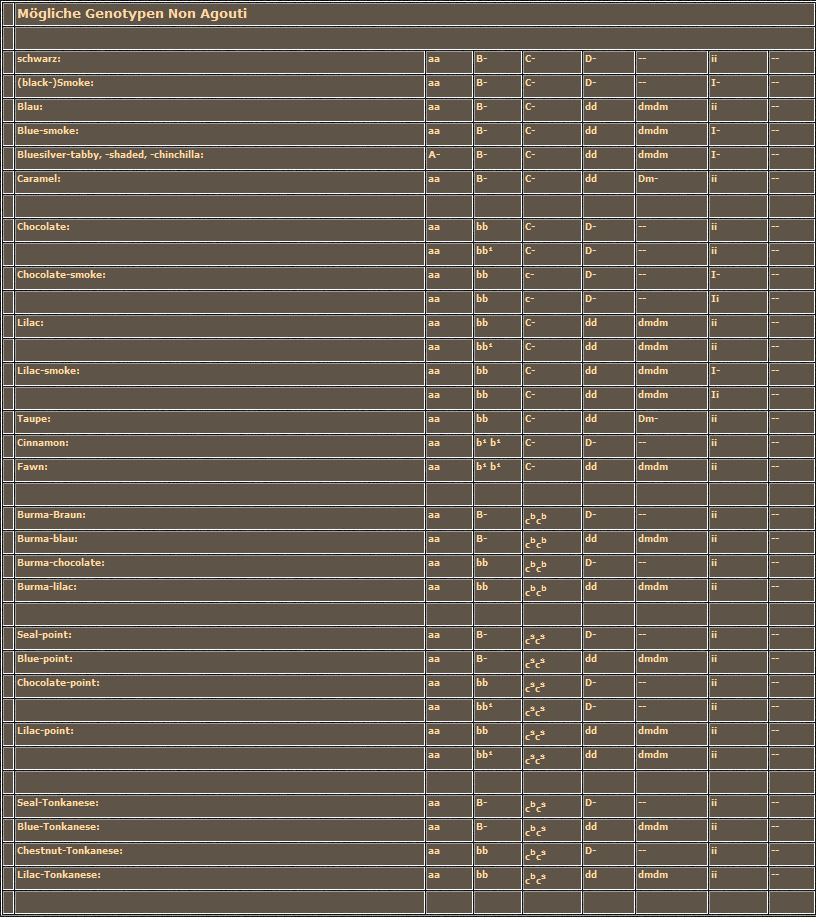
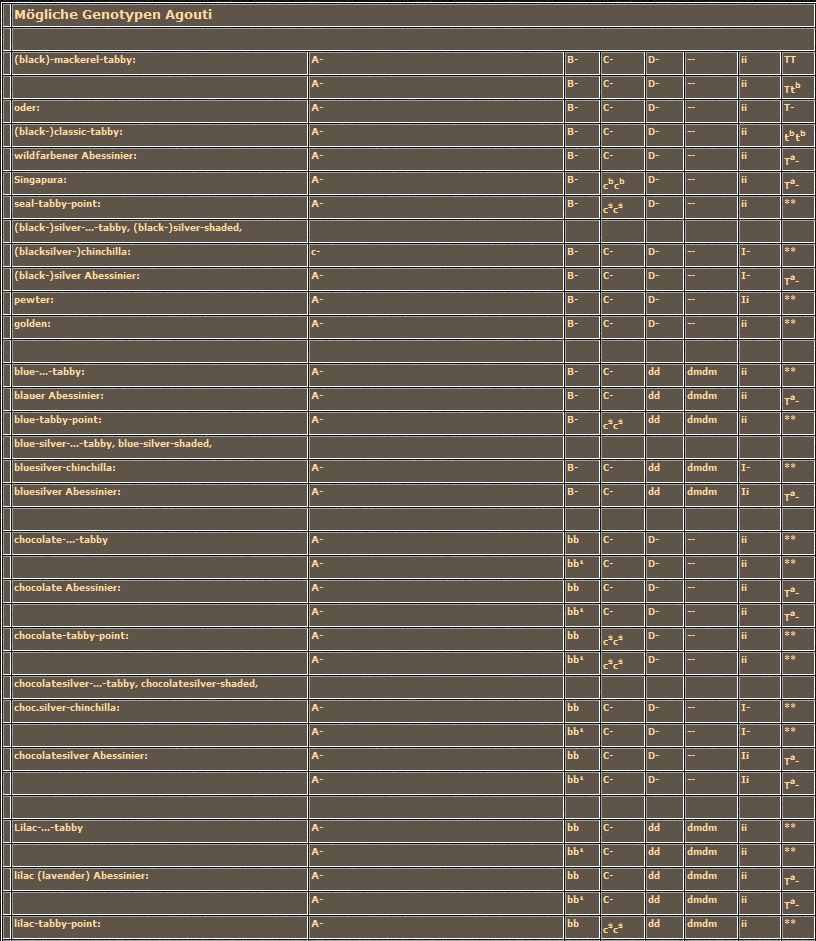
Heute wollen wir uns, wie bereits angekündigt oder fast angedroht, mit den "Silbernen" und den "Roten" auseinandersetzen. Fast angedroht deshalb, weil der Zusammenhang zwischen Genotyp und Phänotyp nicht einfach darzustellen und zu verstehen ist. Die Wirkungen der beiden Farbgene werden von Polygenen so stark beeinflußt, daß gleiche Genotypen bezüglich der Hauptgene zu ganz unterschiedlichen Farbschlägen führen. Außerdem kommen wir nicht umhin, das Gebiet der "einfachen Farben" (Non-Agoutis) zu verlassen. Wir müssen die Tabby-Katzen, die ja alle das Agouti-Allel tragen, ab jetzt mit einbeziehen. Die einzelnen Formen der Tabby-Zeichnungen besprechen wir aber später. Wir merken uns jetzt nur, daß Agouti-Katzen (A/A oder A/a) immer irgendeine Form von Tabby-Zeichnung tragen, während Non-Agouti-Katzen (a/a) an keiner Stelle im Fell ein Tabby-Muster zeigen. Diese Aussage gilt für alle Farben bis auf die "Roten", die wieder einmal die berühmte Ausnahme bilden, die es bekanntlich ja zu jeder Regel geben soll. Rote Katzen oder die roten Flecken bei mehrfarbigen Katzen sind immer tabby-gemustert, egal ob sie Agoutis oder Non-Agoutis sind.
Nach der Besprechung der Gene und deren Allele folgt wieder die obligatorische Tabelle. Ich habe schon einmal erwähnt, daß ich mich darum bemühe, daß die Genotypen-Liste vollständig ist. Allerdings bleiben Genotypen, die zu erheblichen Mißbildungen führen oder die züchterisch nicht von Bedeutung sind, weiterhin außen vor. Sollte ich dennoch etwas vergessen haben oder haben Sie hinsichtlich einer bestimmten Farbe andere Erfahrungen gesammelt, dann tun Sie zweierlei: Üben Sie Nachsicht mit mir und informieren Sie mich, denn auch ich möchte gerne noch etwas dazu lernen.
Das I-Gen (Allele: I, i)
Das Melanin-Inhibitor-Gen wird auch oft Silber-Gen genannt und die Katzen, die das dominante Inhibitor-Allel (I) tragen, der sogenannten "Silber-Gruppe" zugeordnet. Bleiben wir aber lieber bei der Bezeichnung Inhibitor-Gen, denn wie wir gleich sehen werden, sind die wirklich "Silbernen" nur ein kleiner Teil der Katzen mit dem I-Allel.
Allel I = Melanin-Inhibitor
Die Wirkung des mutierten, aber dominanten I-Allels wird oft so beschrieben: Das Pigment (Melanin) wird aus dem Haargrund und dem Unterfell verdrängt. Nur die dunkle Färbung der Haarspitzen, eben der "tips" bleibt erhalten, weshalb man die Gruppe besser als Tipped-Katzen bezeichnen sollte. Die "Silbernen" sind dann einfach eine Untergruppe davon. Und wie wir noch sehen werden, gibt es eine ganze Menge weiterer Untergruppen.
Genauer betrachtet entstehen die Tipped-Haare gerade anders herum. Das Pigment wird nicht im Haar selbst gebildet, sondern in der Haarwurzel, die ja ein Teil der Unterhaut ist. Wenn das Haar wächst, sich also durch die Wurzelscheide nach außen schiebt, wird gleichzeitig Pigment aus der Haarwurzel in das Haar selbst transportiert und dort eingelagert. Das Inhibitor-Gen beeinflußt zusammen mit einer ganzen Reihe anderer Gene diesen Pigment-Transport zum wachsenden Haar. Das I-Allel behindert diesen anfänglich normalen Transport zunehmend. Daher ist die Haarspitze normal gefärbt, während zum Haargrund hin die Farbintensität immer mehr abnimmt. Dadurch, daß dieser Effekt polygen veranlagt ist, kommt es zu einer großen Variationsbreite. Die Tips können unterschiedlich lang sein und die Aufhellung des Haargrundes reicht von einer lediglich leichten Abschwächung der Normalfarbe bis zu einem silberhellen Weiß. Bei den Agoutis bleiben die Haare in der Tabby-Zeichnung fast unbeeinflußt, während die in den dazwischen liegenden Agouti-Bereichen, also die normalerweise typisch gebänderten Haare, maximal aufgehellt sind und nur noch ganz kurze Tips aufweisen. Aufgrund der polygenen Veranlagung kann eine bestimmte Tipped-Variante nur durch konsequente Linienzucht aufrecht erhalten werden.
Smokes ohne Agouti
Fangen wir mit der Beschreibung der Smokes oder Rauchfarbenen an, der einzigen Non-Agouti-Form der Tipped-Katzen. Die Tips machen etwa 2/3 der Haarlänge aus, daher auch bei manchen Autoren die Bezeichnung "starkes Tipping". Der Haargrund ist fast weiß, aber immer noch stärker pigmentiert als in den Agouti-Bereichen der Tabbys. Diese und die folgenden Maßangaben sind nur ungefähre Schätzungen und unterliegen der schon angesprochenen Variationsbreite durch die Polygene. Gehen Sie also bitte nicht bei Ihrer Katze nachmessen, sondern lassen Sie es dort auch bei einer augenscheinlichen Schätzung. Die typischen Smoke-Farben sind die Smoke selbst, besser als Black-Smoke bezeichnet, Blue-Smoke, Chocolate-Smoke und Lilac- bzw. Lavender-Smoke. Eine Smoke-Katze sieht auf den ersten Blick einfarbig aus, erst wenn das Fell geteilt wird sieht man den silberweißen Untergrund. Dieser Effekt ist natürlich bei den dunklen Farben black-smoke, blue-smoke und chocolate-smoke besonders eindrucksvoll. Als Augenfarbe wird bei den Smokes dunkles Gelb bis Kupfer bevorzugt, bei den Orientalisch Kurzhaar sollen die Augen jedoch bei sonst gleichem Genotyp möglichst grün sein. Nasenspiegel und Fußballen tragen die Farbe der Tips.
Tipping nur im Agouti Bereich der Tabbys
Die folgenden Tipped-Varianten sind alle Agoutis. Wenn wir nach der Länge der Tips sortieren, dann sind jetzt die Silbernen dran, die besser Silver-Tabbies genannt werden. Bei ihnen tritt das Tipping nur in den hellen Agouti-Bereichen auf, während die Haare in der Tabby-Zeichnung bis auf einen kurzen hellen Bereich am Haargrund kräftig durchgefärbt sind. Da die Tips in den Agouti-Bereichen nur höchstens 1/3 der Haarlänge ausmachen, ergibt sich ein hoher Kontrast zwischen dem silberweißen Untergrund und dem dunklen Tabby-Muster. Der Nasenspiegel ist rot oder rosa und in der Farbe des Tippings umrandet. Und genau da machen uns die Roten schon wieder einen gewaltigen Strich durch die Rechnung. Red-Smoke und Red-Silver-Tabby (Cameo-Tabby) können den gleichen Genotyp haben, da die Tabby-Zeichnung unabhängig vom Agouti-Allel auftreten kann. Der Kontrast zwischen Grundfarbe und Tabby-Muster kann durch Linienzucht so weit verringert werden, daß die Katze einfarbig mit silberweißem Untergrund erscheint, dann ist es eine Red-Smoke. Ob sie aber Agouti oder Non-Agouti ist, kann man weder bei der Red-Smoke noch bei der Cameo-Tabby mit absoluter Sicherheit erkennen. Da hilft nur ein gründliches Studium der Stammbäume der Elterntiere. Aber bleiben wir bei unseren schon besprochenen Farben. Da steht natürlich an erster Stelle die Farbe Blacksilver-Tabby, die kurzerhand einfach Silver-Tabby genannt wird. Dann folgen, wie bei den Smokes, Bluesilver-Tabby, Chocolatesilver-Tabby und Lilacsilver-Tabby bzw. Lavendersilver-tabby.
Shadeds sind aufgelöste Tabbys
Kommen wir zur nächsten Untergruppe, den Shaded. Die Tips haben etwa dieselbe Länge wie bei den Silver-Tabbies. Entsprechend wird diese Form als "mittleres Tipping" bezeichnet. Auch der Genotyp ist derselbe, es sind Agoutis, die eigentlich eine Tabby-Zeichnung haben müßten. Die Tatsache, daß bei Langhaar-Katzen ein Tabby-Muster in der Regel nicht so deutlich ausfällt und ausdauernde Linienzucht haben dazu geführt, daß die ürsprüngliche Zeichnung sozusagen polygen aufgelöst worden ist. Bei Jungtieren mit kurzem anliegenden Fell kann man als Geisterzeichnung die Herkunft oft noch deutlich erkennen. Auf jeden Fall haben die Shaded einen roten oder rosa Nasenspiegel, der in der Farbe der Tips umrandet ist. Die Augen sind häufig grün. Die Farbbezeichnungen sind ähnlich wie bei den Tabbies, nur daß anstatt -tabby die Endung -shaded angehängt wird. Übersetzt man shaded, kommt "schattiert" heraus. Verwendet man konsequent die deutschen Begriffe, müßte die Silver-Shaded eigentlich schwarzsilber-schattiert heißen. Wenn dann noch ein paar Nuancen dazu kommen, wird schnell ein Zungenbrecher daraus. Bleiben wir also bei den etwas eleganter klingenden englischen Bezeichnungen und definieren die Silver-Shaded als Blacksilver-Shaded.
Wie zu erwarten, sind bei der letzten Gruppe die Tips noch kürzer. Bei dem "leichten Tipping" oder bei den Shells bekennt nur noch 1/8 der Haarlänge Farbe. Zusammen mit dem silberweißen Haargrund verleiht dies dem Fell funkelnden Effekt, der besonders bei Bewegungen deutlich zum Vorschein kommt. Shell kommt von Muschel, wobei sicherlich nicht die Außenseite der Schale gemeint ist, sonder die glitzernde und glänzende Innenseite. Sonst gelten die gleichen Bedingungen wie bei den Shaded. Nur daß bei den Farben schwarz, blau, chocolate und lilac nicht shell, sondern -chinchilla angehängt wird. Steht Chinchilla allein, so ist damit Blacksilver-Chinchilla gemeint. Diese Bezeichnung ist von den gleichnamigen Nagetieren abgeleitet, was allerdings irreführend ist, weil die Farbverteilung genau umgekehrt ist. Die Nager haben einen dunklen Haargrund und helle Spitzen.
Pewter als Zwischenstufe
Um die ganze Sache noch zu verkomplizieren, gibt es noch zwei Ausnahmen, wobei uns da die Genotyp-Analyse völlig im Stich läßt. Wird eine homozygote Silberne (I/I), z.B. Chinchilla oder Shaded, mit einer "nicht-silbernen" Tabby-Katze (i/i) verpaart, sollte daraus nach den Mendel'schen Regeln eine heterozygote Silberne (I,i) entstehen, denn das Inhibitor-Allel ist ja dominant. Nichts dergleichen passiert. Was dabei herauskommt, steht irgendwo zwischen Shaded und Chinchilla. Das Unterfell ist weiß, die üppige Haarspitzenfärbung erweckt den Eindruck, als ob das Tier einen "Farbmantel" trüge, das Tier erscheint zinnfarben, daher der Name Pewter. Der Nasenspiegel ist ziegelrot mit schwarzer Umrandung, die Fußballen sind dunkel und die Augen kupferfarben ohne grünen Einschluß.
Die zweite Ausnahme gehört eigentlich gar nicht zu den Silbernen, weil der Genotyp homozygot nicht-silber (i,i) ist. Es sind Tabby-Katzen, die von heterzygoten (I,i) Chinchillas abstammen. Wohlgemerkt, von echten Chinchillas und nicht von Pewter. Man kann sich diesen besonderen Farbschlag nur dadurch erklären, daß die Polygene der Chinchilla-Eltern trotz Nicht-Silber-Genotyp voll wirksam sind. Der erfahrene Züchter weiß schon, worauf ich hinaus will, auf das zweite Edelmetall, die Goldenen oder Golden-Tabbies. Der Haargrund ist nicht weiß, sondern zeigt ein intensives warmes Creme. Das kommt daher, daß in den Agouti-Bereichen das gelbe Agouti-Band der Haare ausgeweitet ist, die Polygene der Eltern verstärken die Gelb-Pigmentierung. Erinnern wir uns, die normale Agouti-Bänderung ist heller Haargrund, dunkles Band, gelbes Agouti-Band und dunkle Haarspitze. Bei den Golden-Tabbies ist der Haargrund hellblau, dann folgt das ausgeweitete intensivgelbe Agouti-Band und zuletzt die in der Regel schwarze Haarspitze. Aber auch andere Farben werden schon gezüchtet, so gibt es insbesondere bei den Kurzhaar-Rassen schon chocolate-golden, blue-golden und vereinzelt lilac-golden. Bei den ursprünglichen Golden-Tabbies ist der Nasenspiegel tiefrosa mit sealbrauner Umrandung, die Fußballen sind ebenfalls sealbraun, die Augen grün, höchstens blau-grün.
Allel i = normaler Melanineinbau
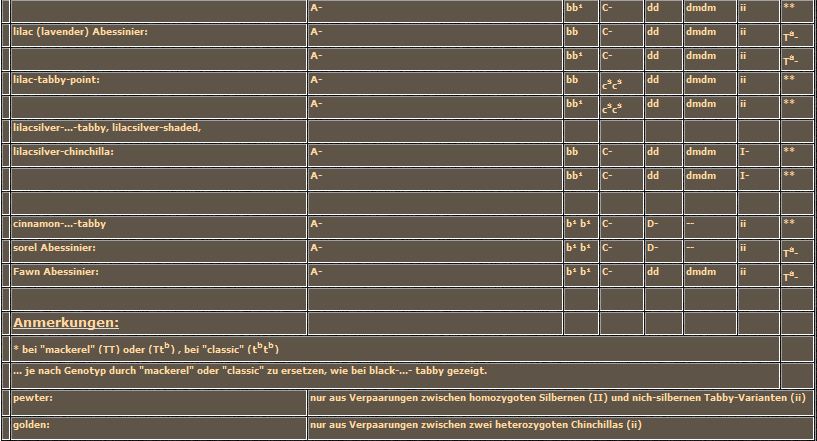
Die Wirkung des in diesem Fall rezessiven Wildtyp-Allels ist auf jeden Fall schneller beschrieben als die des mutierten Allels. Die normale Farbstoffeinlagerung in das wachsende Haar wird nicht behindert. Es entstehen auf der ganzen Länge durchgefärbte Haare oder in den Agouti-Bereichen Haare mit der gewohnten Agouti-Bänderung. Ich habe in diesem Abschnitt den Begriff Melanin absichtlich durch die allgemeine Bezeichnung Farbstoff ersetzt, weil der fälschlicherweise als "Melanin-Inhibitor" bekannte Faktor natürlich nicht nur auf das Melanin wirkt, sondern auf alle uns bekannten Fellfarb-Pigmente. Aber diese Tatsache ist Ihnen sicher schon bei der Besprechung von chocolate-silver aufgefallen. Auch wenn wir die "Roten" besprechen, werden wir den Melanin-Inhibitor nochmal aus der Schublade ziehen müssen, auch wenn die Farbe rot oder orange nichts mit Melanin zu tun hat.
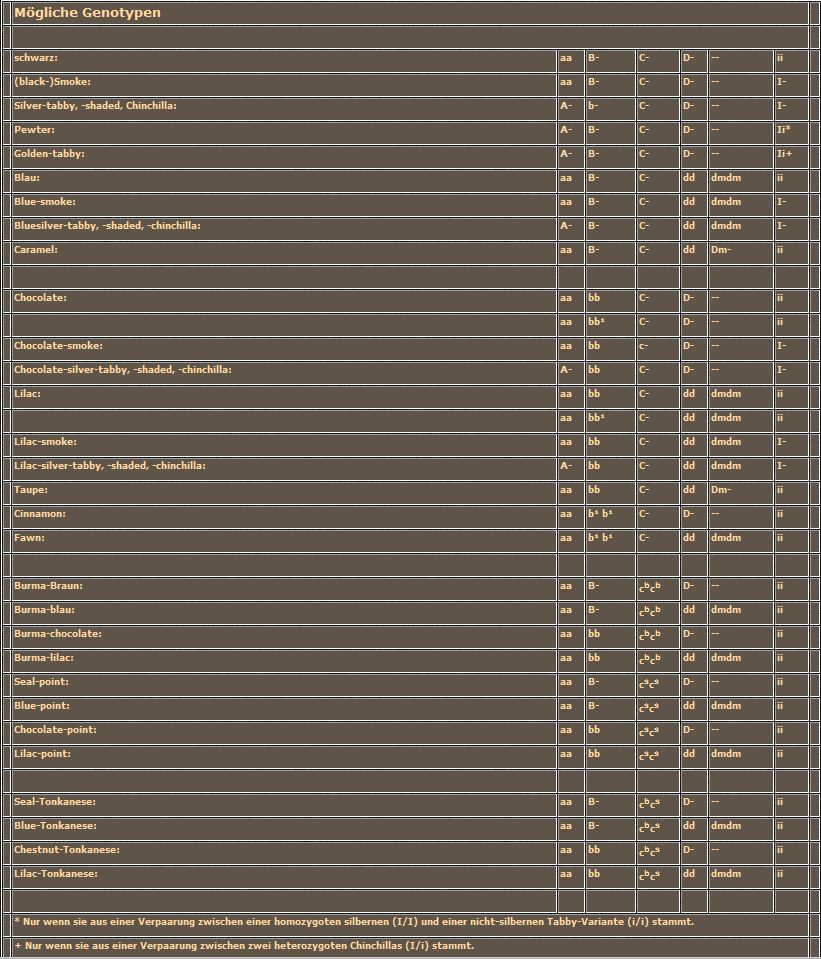
Jetzt ist wieder einmal ein Tabelle fällig. Wie Sie sehen, habe ich den Dm-Faktor doch noch einmal mitgeschleppt. Nur damit Sie sich erinnern, daß unsere Katzen normalerweise die homozygot-rezessive Allelenkombination tragen, lediglich für blau und lilac führt die weitere Modifikation der Verdünnung zu ansprechbaren Ergebnissen. Neu ist das Agouti-Allel und der (Melanin)-Inhibitor mit seinen beiden Schalterstellungen.
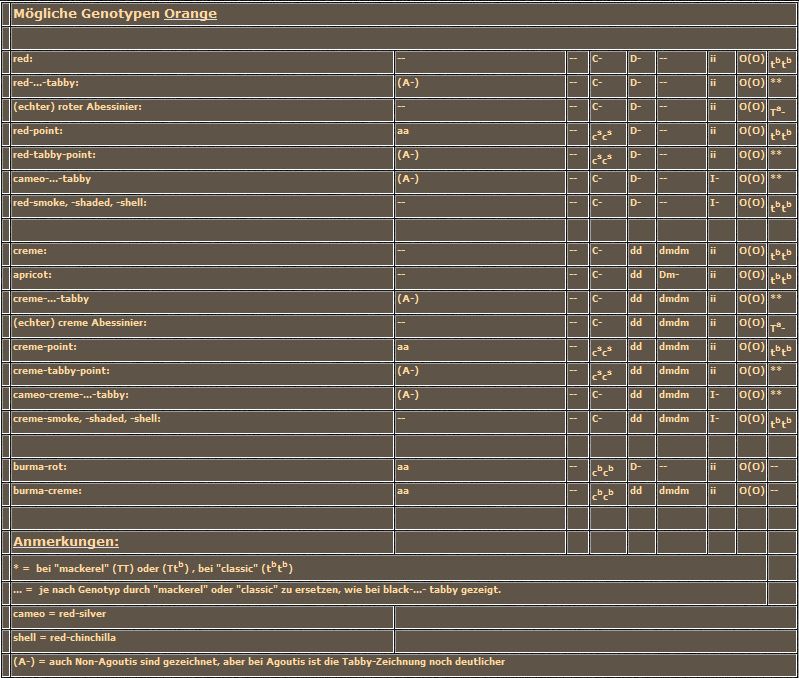


 wiederholt sich nur in den Katern. Neu ist das Orange-Gen in heterozygoter Allelenausstattung (Ox, ox), die als Schildpatt bezeichnet wird. Wie diese besondere Zeichnung zustande kommt ohne daß irgendwelche Dominanz-Rezessivitätsbeziehungen vorliegen, das besprechen wir gleich im Anschluß. Machen wir zuerst die reziproke Kreuzung. Die "rote" Katze produziert wieder nur eine Sorte von Eizellen, nämlich solche mit dem orange-Allel (Ox) auf den X-Chromosomen. Die Katze muß ja homozygot orange sein, sonst wäre sie ja schildpatt. Der "schwarze" Kater hingegen liefert zwei Spermientypen, die mit dem nicht-orange tragenden X-Chromosom (ox) und die mit dem y-Chromosom.
wiederholt sich nur in den Katern. Neu ist das Orange-Gen in heterozygoter Allelenausstattung (Ox, ox), die als Schildpatt bezeichnet wird. Wie diese besondere Zeichnung zustande kommt ohne daß irgendwelche Dominanz-Rezessivitätsbeziehungen vorliegen, das besprechen wir gleich im Anschluß. Machen wir zuerst die reziproke Kreuzung. Die "rote" Katze produziert wieder nur eine Sorte von Eizellen, nämlich solche mit dem orange-Allel (Ox) auf den X-Chromosomen. Die Katze muß ja homozygot orange sein, sonst wäre sie ja schildpatt. Der "schwarze" Kater hingegen liefert zwei Spermientypen, die mit dem nicht-orange tragenden X-Chromosom (ox) und die mit dem y-Chromosom. Nehmen wir uns zuerst die unterste Reihe vor. Sowohl in der ersten als auch in der zweiten Kreuzung ist die eine Hälfte der Kater "schwarz", die andere "rot". Bei den Mädchen in der oberen Spalte sieht eine Hälfte immer so aus wie der Vater, also "schwarz" aus der ersten Kreuzung und "rot" aus der zweiten. Nur die zweite Hälfte der Mädchen oder insgesamt 1/4 der gesamten Nachkommen sind schildpatt. Aber glauben Sie ja nicht, damit einen Weg zur Reinzucht von Schildpatt gefunden zu haben. Durch die immer wieder notwendige Einkreuzung von nicht-schildpatt Katern werden bei jeder Verpaarung laufend eine ganze Reihe von Faktoren, die die Ausprägung des Schildpatt-Musters mitbestimmen, neu kombiniert. Damit haben Sie als Züchter zwar die Möglichkeit vorauszusagen, aus welcher Verpaarung mit welcher Wahrscheinlichkeit (25% bis höchstens 50%) Schildpatt-Katzen fallen werden, aber die Farbverteilung wird sich immer Ihrer Kontrolle entziehen. Dies gilt auch dann, wenn Sie immer wieder die gleiche Katze mit dem gleichen Kater verpaaren, dazu gleich mehr.
Nehmen wir uns zuerst die unterste Reihe vor. Sowohl in der ersten als auch in der zweiten Kreuzung ist die eine Hälfte der Kater "schwarz", die andere "rot". Bei den Mädchen in der oberen Spalte sieht eine Hälfte immer so aus wie der Vater, also "schwarz" aus der ersten Kreuzung und "rot" aus der zweiten. Nur die zweite Hälfte der Mädchen oder insgesamt 1/4 der gesamten Nachkommen sind schildpatt. Aber glauben Sie ja nicht, damit einen Weg zur Reinzucht von Schildpatt gefunden zu haben. Durch die immer wieder notwendige Einkreuzung von nicht-schildpatt Katern werden bei jeder Verpaarung laufend eine ganze Reihe von Faktoren, die die Ausprägung des Schildpatt-Musters mitbestimmen, neu kombiniert. Damit haben Sie als Züchter zwar die Möglichkeit vorauszusagen, aus welcher Verpaarung mit welcher Wahrscheinlichkeit (25% bis höchstens 50%) Schildpatt-Katzen fallen werden, aber die Farbverteilung wird sich immer Ihrer Kontrolle entziehen. Dies gilt auch dann, wenn Sie immer wieder die gleiche Katze mit dem gleichen Kater verpaaren, dazu gleich mehr.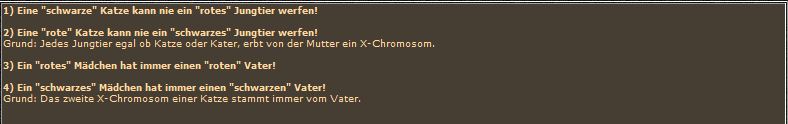
 In einem ganz bestimmten Stadium formt sich der Zellhaufen zu einer Hohlkugel um, dann gehen die mitotischen Teilungen weiter. Dabei werden ganz bestimmte Einfaltungen vorgenommen, bestimmte Bereiche differenzieren sich während der weiteren Teilungen zu ganz bestimmten Organen wie Nervensystem, Skelett, innere Organe, Haut, Muskeln etc.. Der Differenzierungszustand wird nun bei jeder weiteren Teilung an die Tochterzellen weitergegeben, so daß z.B. aus Hautzellen immer wieder nur Hautzellen entstehen. Irgendwann, nach vielen Mitosen, ist dann die kleine Katze fertig und kommt auf die Welt. Weitere Zellteilung lassen das Tier heranwachsen und wieder gilt der Grundsatz, daß der einmal festgelegte Differenzierungszustand bei jeder Teilung unverändert an die Tochterzellen weitergegeben wird. Ist die Katze erwachsen, werden die Teilungszyklen langsamer und seltener, denn es müssen jetzt nur noch verbrauchte, defekte oder abgestorbene Zellen ersetzt werden. Nach einer bestimmten Zeitspanne, der Lebenszeit, die bei der Katze 15-20 Jahre dauert, beginnen die Mitosen ungenau zu werden. Das Erbmaterial wird nicht mehr korrekt auf die Tochterzellen verteilt, der Differenzierungszustand wird unvollständig weitergegeben oder geht ganz verloren. Die Organe funktionieren dann nicht mehr richtig und wenn wichtige Organe ganz ausfallen, dann stirbt die Katze. Das ist sehr schlicht ausgedrückt der Lebenszyklus aus der Sicht der Genetik und der Zellbiologie.
In einem ganz bestimmten Stadium formt sich der Zellhaufen zu einer Hohlkugel um, dann gehen die mitotischen Teilungen weiter. Dabei werden ganz bestimmte Einfaltungen vorgenommen, bestimmte Bereiche differenzieren sich während der weiteren Teilungen zu ganz bestimmten Organen wie Nervensystem, Skelett, innere Organe, Haut, Muskeln etc.. Der Differenzierungszustand wird nun bei jeder weiteren Teilung an die Tochterzellen weitergegeben, so daß z.B. aus Hautzellen immer wieder nur Hautzellen entstehen. Irgendwann, nach vielen Mitosen, ist dann die kleine Katze fertig und kommt auf die Welt. Weitere Zellteilung lassen das Tier heranwachsen und wieder gilt der Grundsatz, daß der einmal festgelegte Differenzierungszustand bei jeder Teilung unverändert an die Tochterzellen weitergegeben wird. Ist die Katze erwachsen, werden die Teilungszyklen langsamer und seltener, denn es müssen jetzt nur noch verbrauchte, defekte oder abgestorbene Zellen ersetzt werden. Nach einer bestimmten Zeitspanne, der Lebenszeit, die bei der Katze 15-20 Jahre dauert, beginnen die Mitosen ungenau zu werden. Das Erbmaterial wird nicht mehr korrekt auf die Tochterzellen verteilt, der Differenzierungszustand wird unvollständig weitergegeben oder geht ganz verloren. Die Organe funktionieren dann nicht mehr richtig und wenn wichtige Organe ganz ausfallen, dann stirbt die Katze. Das ist sehr schlicht ausgedrückt der Lebenszyklus aus der Sicht der Genetik und der Zellbiologie.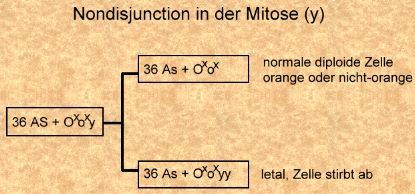


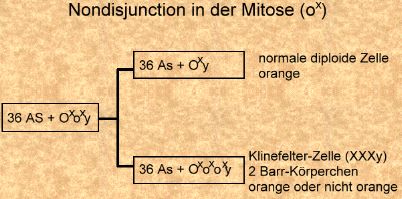
 Nondisjunction in der Meiose I und Befruchtung mit normalen Spermien eines nicht-orange Katers (Bild rechts).
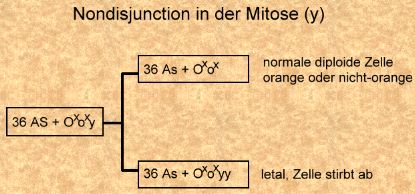
Nondisjunction in der Meiose I und Befruchtung mit normalen Spermien eines nicht-orange Katers (Bild rechts). Nondisjunction in der Meiose I und Befruchtung mit normalen Spermien eines orangen Katers (Bild links).
Nondisjunction in der Meiose I und Befruchtung mit normalen Spermien eines orangen Katers (Bild links).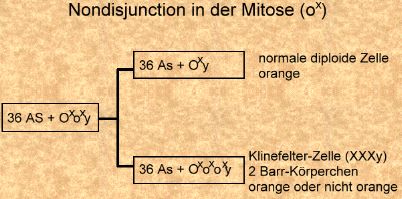
 Bei den Bicolor- und Tricolor-Katzen soll mindestens ein Drittel und höchstens die Hälfte der Körper-fläche weiß sein (Grad 3 bis Grad 5).
Bei den Bicolor- und Tricolor-Katzen soll mindestens ein Drittel und höchstens die Hälfte der Körper-fläche weiß sein (Grad 3 bis Grad 5).