Teil 1
Die Vererbungslehre ist eine recht junge Wissenschaft und, was den praktischen Nutzen der Vererbungsgesetzte angeht, eine recht undankbare dazu. Jeder Hobbyzüchter hat sicher schon einmal die folgende Situation erlebt: Man hat Berge von Kreuzungstabellen und Büchern gewälzt, mit dem so erworbenen Wissen eine tolle Verpaarung geplant, aufgrund der theoretischen Recherchen, mit einer Vielzahl von außergewöhnlichen Farbvarianten gerechnet und dann voller Erwartung vor der Wurfkiste gesessen. Und was machte die liebe Katzenmama? Sie brachte völlig selbstverständlich lauter muntere liebe Katzenkinder zur Welt, aber alle gleichfarbig und vielleicht auch noch lauter Katerchen oder Kätzchen. Dabei weiß doch jeder, auch der, der sich noch nie in seinem Leben um Vererbungsgesetze gekümmert hat, daß normalerweise immer etwas gleichviel Kater und Katzen zu erwarten sind. Ist also die ganze Vererbungslehre wirklich nur Humbug? Oder steckt hinter diesem Zufall doch ein System?
Vererbung funktioniert nach Gesetzmäßigkeiten
Also „Humbug" ist die Vererbungslehre oder Genetik, wie sie seit dem Jahr 1906 heißt, sicher nicht. Dann bleibt uns nur übrig, die Gesetzmäßigkeiten zu finden, die hinter all den Zufällen stecken. Dabei wird es uns nicht erspart bleiben, neben dem Begriff Genetik auch noch ein paar andere Fachbegriffe zu erarbeiten. Ich will damit niemanden ärgern oder langweilen, aber Fachwörter stehen immer für genau definierte komplizierte Sachverhalte. Sie geben uns die Möglichkeit, vielschichtige, umfangreiche Probleme kurz, präzise und unmißverständlich darzustellen. Dies gelingt jedoch nur, wenn alle die einzelnen Fachwörter gleich verwenden. Wenn wir uns mit Vererbungslehre oder Genetik beschäftigen, dann beobachten wir, wie einzelne Merkmale von einer Generation an die andere weitergegeben werden oder wie sich bestimmte Merkmale von Katze oder Kater in den Babys wiederfinden, vermischen oder verstecken, also scheinbar verlorengehen. Aber schon der Begriff Merkmal führt leicht zu Mißverständnissen. Was ist ein Merkmal? Nehmen wir das Fell unserer Kuscheltiere als Beispiel. Ist vielleicht „Kurzhaar blau" ein Merkmal? Oder nur „Fell", wobei dann „kurzes Haar" und „blau" enthalten sind? Oder sind etwa „Fellänge" und „Fellfarbe" zwei getrennte Merkmale? Um hier Begriffe exakt zu definieren, müssen wir etwas tiefer in die Trickkiste der Natur greifen.
Baupläne als Informationsträger
Das Wesen der Vererbung besteht darin, daß ein bestimmtes Erbgut, also ein bestimmter Bauplan, von Generation zu Generation weitergegeben wird. Die Summe aller in einem Bauplan für einen bestimmten Organismus enthaltenen Informationen nenne wir Genom. Bei einer Katze enthält die befruchtete Eizelle alle die Einzelinformationen als Genom zusammengefaßt, die dann während der Entwicklung alle die Merkmale ergeben, an denen wir diesen Organismus als Katze erkennen. Und was gehört so alles zu einer Katze? Ein bestimmter Körperbau, Fellqualität, eine bestimmte Kopfform, Ohrenform, Augenform, Augenfarbe, Fellfarbe, Fellänge, Farbdichte . . . ! Halt! Hier
hat sich von ganz allein eine Begriffsbestimmung ergeben. Wenn das Genom die Informationen für alle Merkmale enthält, dann enthält jedes Gen, das ja Teil des Genoms ist, die Information für ein Merkmal. Dabei sind manche Merkmale offensichtlich, wie zum Beispiel Fellfarbe, Fellänge oder Farbdichte, die meisten aber, wie zum Beispiel Körperbau, innere Organe etc., eher versteckt und für unsere Zwecke auch nicht so wichtig. Und was ist nun mit „kurzem Haar", „langem Haar" oder „schwarzem Fell", „blauem Fell"? Nun, hier handelt es sich um Ausprägungen oder Schalterstellungen eines bestimmten Merkmals oder Gens, wir nennen dieses in Zukunft Allel. Also nochmals: Jedem Merkmal, zum Beispiel „Farbdichte", liegt ein bestimmtes Gen zugrunde, daß alle notwendigen Informationen zur Ausbildung des Merkmals enthält, hier in unserem Beispiel also die „Form und Verteilung" der Pigmente steuert, die die Fellfarbe entstehen läßt. Das Gen kann
aber auch noch verschiedene Schalterstellungen oder Allele einnehmen, die dann zu der „vollen" Farbe Schwarz oder der „verdünnten" Farbe Blau zu führen.
Mendelsche Gesetze sind bin heute Grundlage
Wo sind nun diese Gene, die statistisch gesehen nach bestimmten Regeln, im Einzelfall jedoch mehr zufällig von den Eltern auf die Kinder weitergeben werden? Die gedankliche Grundlage lieferte 1866 der Augustinermönch Johann Gregor Mendel, der eine riesige Zahl von Erbsen kreuzte und die Nachkommenschaft statistisch auswertete. Er leitete daraus die heute noch gültigen Mendel'schcn Regeln der Vererbung ab, mit denen wir uns später noch genauer beschäftigen werden. Er fand aber
auch heraus, daß die meisten Gene im Prinzip unabhängig voneinander weitergegeben werden, manche jedoch fast immer aneinander gekoppelt auftreten. Im Zellkern einer jeden Zelle, auch der Eizelle, aus der jeder Organismus durch eine Anzahl von Zellteilungen, den Mitosen, schließlich entsteht, fand man anfärbbare Körperchen, die Chromosomen. Diese Chromosomen machen während der Zellteilungen bestimmte Zyklen durch und werden nach bestimmten Regeln auf die beiden Teilungsprodukte, die Tochterzellen, verteilt. Die Anzahl und Form der Chromosomen ist in jeder Zelle einer Pflanzen- oder Tierart immer gleich und in Zellen nahe verwandter Arten ähnlich. Es
wurde schon 1885 geschlossen, daß die Chromosomen die Träger der Gene sind. Gene, die unabhängig voneinander vererbt werden, liegen auf verschiedenen Chromosomen, solche die aneinander gekoppelt vererbt werden, liegen hintereinander auf demselben Chromosom. Noch interessanter sind die speziellen Teilungen, die zu den Keimzellen oder Gameten führen, also bei der Katze zu den Eizellen und beim Kater zu den Spermien. Diese Reifeteilungen oder Meiosen sorgen dafür, daß das Erbgut von Mutter und Vater nach bestimmten Regeln, auf die wir später noch
genauer eingehen wollen, auf die Eizellen und Spermien verteilt wird. Nach dem Decken verschmelzen je eine Eizelle und ein Spermium zur befruchteten Eizelle, der Zygote, aus der dann durch Mitosen wieder ein Kätzchen entsteht. Wenn wir also den Weg bestimmter Chromosomen gedanklich über mehrere Generationen verfolgen und dabei die Verteilungsregeln in den Teilungszyklen der Meiosen beachten, müßten wir die Mendel'schen Regeln nachvollziehen können und damit verstehen lernen, welches System hinter dem Zufall „Vererbung" steckt.
Die junge Wissenschaft kämpft mit einigen Unsicherheiten
Bevor wir uns an die zugegebenermaßen etwas schwierige Aufgabe machen, die Grundlagen der Vererbungsgesetze zu verarbeiten, ein kleiner historischer Seitenblick. Die Geschichte ist nicht nur interessant, sie zeigt uns auch, wie jung die Vererbungslehre noch ist. Und da die Vererbungsregeln nur statistisch zu erfassen sind, ist es nur verständlich, daß es bei dem kurzen Erfahrungszeitraum noch manche Unsicherheiten gibt. Dazu kommt noch eine Schwierigkeit, vor der wir heute mit unseren ganzen modernen Methoden genauso stehen wie Mendel vor 125 Jahren. Wir können genausowenig wie Mendel ein Gen sehen oder irgendwie kenntlich machen. Erst wenn von einem
Gen zwei oder mehr Allele existieren, erkennen wir es als eigenständiges Gen. Wenn es zum Beispiel nur Katzen mit kurzem Fell gäbe, bestünde kein Grund anzunehmen, daß es ein eigenes Gen gibt, das für das Merkmal „Fellänge" zuständig ist. Erst wenn durch eine sprunghafte Änderung der genetischen Information, durch eine Mutation, eine vom Wildtyp oder Normaltyp abweichende Ausprägung eines Merkmals, also ein neues Allel entstanden ist, können wir daraus schließen, daß es
sich um ein selbständiges Gen handelt. Wie funktioniert nun so eine Mutation? Daß die Gene auf den Chromosomen liegen, haben wir schon gesehen.
Mutationen sind Sprunghafte Zufälligkeiten
Die Chromosomen bestehen aus Eiweißen oder Proteinen und den Nucleinsäuren. Diese ominösen Säuren heißen so, weil sie nur im Zellkern, dem Nucleus, vorkommen. Die wichtigsten Nucleinsäuren sind die Desoxyribonucleinsäure und die Ribonucleinsäure, besser bekannt unter den Kürzeln DNS und RNS. Für den Bauplan eines Lebewesens ist eine fast unendlich große Zahl von Informationen nötig, und diese Information ist in einer speziellen Sprache auf den Chromosomen gespeichert. Unser Alphabet besteht aus 26 Buchstaben, die in einer bestimmten Reihenfolge hintereinander aufgereiht eine bestimmte Information vermitteln. Die Proteine und die Nucleinsäuren sind chemische Verbindungen, die jeweils aus einer bestimmten Anzahl von Grundbausteinen bestehen, die wie bei einer Perlenschnur aufgereiht sind. Die Proteine bestehen aus einer Perlenschnur von Aminosäuren, und es gibt davon 20 Sorten. Die Sache erinnert also stark an unser Alphabet. Die Nucleinsäuren bestehen aus einer Perlenschnur von Nucleotiden und davon gibt es ganze vier Sorten. So schien es klar, daß als Träger des umfangreichen Genoms eines Lebewesens nur die Proteine in Frage kommen. Es sollte noch bis 1944 dauern, bis bewiesen werden konnte, daß nicht die Proteine, sondern entgegen allen Zweifeln die Nucleinsäuren die Träger der Erbinformation sind. 1945 lag dann endlich auch genug statistisch auswertbares Untersuchungsmaterial vor, mit dessen Hilfe gezeigt werden konnte, daß die Mendel'schen Regeln auch bei Tieren anzuwenden sind. 1953 wurde die Struktur der Nucleinsäuren beschrieben. Die beiden Forscher Watson und Crick sollten dafür den Nobel-Preis bekommen. Der genetische Code, das Alphabet, in dem die genetische Information durch die Nucleinsäuren auf die Chromosomen geschrieben ist, wurde schließlich 1966 geknackt. Sie sehen also, 1866 fand Mendel, daß etwas geschrieben steht über den Bauplan eines
jeden Lebenwesens. Aber es dauerte 100 Jahre, bis man beweisen konnte, was und wo und wie die genetische Information gespeichert ist und wie die Baupläne von Generation zu Generation weitergegeben werden.
Genetischer Code auf vier Buchstaben
Lassen Sie uns das Pferd quasi von hinten aufzäumen. Nachdem wir uns heute das Wesen der Mutation anhand des genetischen Codes klargemacht haben, werden wir versuchen, anhand der Geschehnisse bei der Meiose, der Verteilungsregeln des Erbgutes auf die Keimzellen der Meiose, der Verteilungsregeln des Erbgutes auf die Keimzellen und der Befruchtung, die Mendel'schen Regeln zu durchschauen. Dann können wir Kreuzungstabellen lesen, und was noch wichtiger ist, auch interpretieren. Dann können uns solche Zufälle wie der am Anfang beschriebene nicht
mehr schocken, weil wir wissen, welches statistische System dahinter steckt. Nun zum genetischen Code. Wir haben schon gesehen, daß die DNS aus vier Bausteinen, den Nucleotiden besteht. Diese heißen Adenin, Guanin, Thymin und Cytosin. Sie unterscheiden sich chemisch voneinander und zeigen unterschiedliche Eigenschaften. Die Unterschiede sind zwar deutlich, chemisch gesehen jedoch so klein, daß durch äußere Einflüsse, aber auch spontan, das heißt ohne äußeren Anstoß, das eine Nucleotid in ein anderes Nucleotid oder in ein außergewöhnliches Nucleotid mit anderen Eigenschaften umgewandelt werden kann. Machen wir uns ein einfaches Modell, um diese hochtheoretischen Erkenntnisse zu durchleuchten. Wenn wir die einzelnen Nucleotide durch ihre Anfangsbuchstaben bezeichnen, dann könnte ein Ausschnitt aus einem Nucleotidstrang so aussehen:
 Eigenschaften der Proteine durch Reihenfolge der Aminosäuren bestimmt
Eigenschaften der Proteine durch Reihenfolge der Aminosäuren bestimmt
Das soll eine Information sein? Nun gut, spinnen wir die Idee weiter. Was soll der genetische Code eigentlich verschlüsseln? Ein Lebewesen ist aus einer Menge von unterschiedlichen Proteinen aufgebaut. Die Unterschiede in den Proteinen kommen von unterschiedlichen Abfolgen der sie bildenden Aminosäuren, die Reihenfolge macht, daß aus dem einen Protein Teil einer Muskelfaser, aus einem anderen Teil einer Sehne und noch aus einem anderen ein Enzym wird. das zum Beispiel einen Stoffwechselvorgang steuert. Die Liste der Beispiele ließe sich noch fast endlos fortsetzen. Wichtig ist die Erkenntnis, daß die Eigenschaften der Proteine durch die Reihenfolge der Aminosäuren bestimmt werden und daß der genetische Code durch die Reihenfolge der Nucleotide die Baupläne für die verschiedenen Proteine eines Lebenwesens liefern muß. Der genetische Code muß also so beschaffen sein, daß die Reihenfolge der vier Nucleotide mindestens 20 Buchstaben ergibt, für jede Aminosäure einen. Das erinnert stark an das Morsealphabet, wo mit zwei Zeichen, dem Punkt und dem Strich, 26 Buchstaben codiert werden, die Symbole für die einzelnen Buchstaben sind durch Leerzeichen voneinander getrennt. Die DNS-Perlschnüre sind lückenlos, es gibt kein Leerzeichen, dafür aber bestimmte Steuerzeichen wie „Anfang" und „Ende". Mit unseren vier Elementen müssen mehr als 20 Buchstaben dargestellt werden. Wenn wir immer eine Zweiergruppe zu einem Wort zusammenfassen, ergibt das 16 (vier hoch zwei) Möglichkeiten der Kombination, also 16 Buchstaben. Fassen wir immer drei zusammen, dann haben wir 64 (4 hoch 3)
Kombinationsmöglichkeiten. Da wir mindestens 20, mit Anfang und Ende mindestens 24 Buchstaben brauchen, ist das die Lösung. Tatsächlich stellt immer eine Dreiergruppe ein sogenanntes Triplett von
Nucleotiden, einen Buchstaben des genetischen Codes dar. Jedes Triplett steht für eine Aminosäure oder einen Steuerbefehl. Bauen wir an unserem Modell weiter und legen wir die Bedeutung einiger Tripletts fest.

Nehmen wir nochmal unsere erste Nucleotidfolge zur Hand. Jetzt sehen wir, daß in diesem scheinbaren Chaos eine Information steckt.
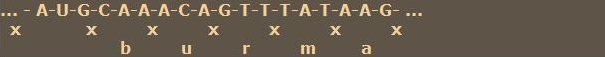
Jetzt lassen wir unser Modell mutieren, entweder spontan oder durch einen äußeren Einfluß wie radioaktive Strahlung, Röntgenstrahlen, UV-Strahlen oder eine giftige chemische Substanz. Nehmen wir an, aus dem C, das genau über dem „u" von „burma" steht, wird ein T.

Mit diesem kleinen Modell haben wir schon das Wesen der Mutation erkannt: eine kleine, punktuelle Veränderung im Genom kann zu deutlich sichtbaren Veränderungen in der Ausprägung eines Merkmals führen. Ein Alle! ist entstanden. Nachdem wir nun wissen, wie ein Gen im Prinzip aufgebaut ist, wie ein Merkmal im genetischen Code verschlüsselt ist, wie sich die Ausprägung eines Merkmals verändern kann, werden wir das nächste Mal erfahren, wie die Merkmale und deren Ausprägungen von Generation zu Generation weitervererbt werden. Mitose und Meiose, die zufällige Verteilung der Allele auf die Keimzellen und die zufällige Kombination der Allele durch die Verschmelzung der Keimzellen zur befruchteten Eizelle werden dann unser Thema sein.




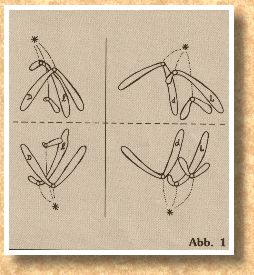
 Jedes Chromosom hat eine "Kupplung", an der später eine Art "Zugseil", die Spindelfaser, angeheftet wird, mit deren Hilfe die Chromosomen auf die beiden Tochterzellen verteilt werden. Diese Stelle wird Zentromer genannt und ist im Schema durch einen Kreis gekennzeichnet. Die Lage des Zentromers ist bei bestimmten Chromosom immer gleich, entweder mehr in der Mitte, zu einem Ende hin verschoben oder fast ganz an einem Ende. Außerdem sehen wir in Abb. 1 und Abb. 2, daß die Chromosomen unterschiedlich groß sein können. Jetzt haben wir schon zwei Unterscheidungsmerkmale, mit deren Hilfe wir Homologenpaare suchen können. Es ergibt sich zwangsläufig die PaarAnordnung wie in Abb. 3 mit drei Homologenpaaren.
Jedes Chromosom hat eine "Kupplung", an der später eine Art "Zugseil", die Spindelfaser, angeheftet wird, mit deren Hilfe die Chromosomen auf die beiden Tochterzellen verteilt werden. Diese Stelle wird Zentromer genannt und ist im Schema durch einen Kreis gekennzeichnet. Die Lage des Zentromers ist bei bestimmten Chromosom immer gleich, entweder mehr in der Mitte, zu einem Ende hin verschoben oder fast ganz an einem Ende. Außerdem sehen wir in Abb. 1 und Abb. 2, daß die Chromosomen unterschiedlich groß sein können. Jetzt haben wir schon zwei Unterscheidungsmerkmale, mit deren Hilfe wir Homologenpaare suchen können. Es ergibt sich zwangsläufig die PaarAnordnung wie in Abb. 3 mit drei Homologenpaaren.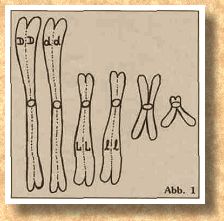 Was sollen die Buchstaben auf zwei von den drei Homologenpaaren? Wir wissen inzwischen, daß die Chromosomen die Träger der Gene sind und daß die Abfolge der Nucleotide auf dem DNS-Strang die Information für ein Merkmal codiert. Mit unserer bereits genetisch geübten Denkweise schließen wir, daß homologe Chromosomen so gleich aussehen, weil sie auch gleiche Gene tragen, und liegen damit absolut richtig. Allerdings können die beiden sich entsprechenden Gene auf den homologen Chromosomen durchaus verschiedene Allele codieren.
Was sollen die Buchstaben auf zwei von den drei Homologenpaaren? Wir wissen inzwischen, daß die Chromosomen die Träger der Gene sind und daß die Abfolge der Nucleotide auf dem DNS-Strang die Information für ein Merkmal codiert. Mit unserer bereits genetisch geübten Denkweise schließen wir, daß homologe Chromosomen so gleich aussehen, weil sie auch gleiche Gene tragen, und liegen damit absolut richtig. Allerdings können die beiden sich entsprechenden Gene auf den homologen Chromosomen durchaus verschiedene Allele codieren. Die männlichen Geschlechtsrealisatoren befinden sich auf den Autosomen. Ist nur ein X-Chromosom vorhanden, entwickelt sich ein männliches Tier, zwei X-Chromosomen führen zu einem Weibchen. Die Schmetterlinge zeigen eine besonders pikante Variante: Die Weibchen haben den XY-Genotyp und die Männchen haben zwei X-Chromosomen. Bei der Katze hat sich jedoch, wie bei allen Wirbeltieren, im Verlauf der Evolution die XY Geschlechtsbestimmung durchgesetzt. Alle Tiere ohne Y-Chromosom sind weiblich, alle Tiere mit mindestens einem Y-Chromosom sind männlich. Also sind im Normalfall XX-Katzen wie in Ab-
Die männlichen Geschlechtsrealisatoren befinden sich auf den Autosomen. Ist nur ein X-Chromosom vorhanden, entwickelt sich ein männliches Tier, zwei X-Chromosomen führen zu einem Weibchen. Die Schmetterlinge zeigen eine besonders pikante Variante: Die Weibchen haben den XY-Genotyp und die Männchen haben zwei X-Chromosomen. Bei der Katze hat sich jedoch, wie bei allen Wirbeltieren, im Verlauf der Evolution die XY Geschlechtsbestimmung durchgesetzt. Alle Tiere ohne Y-Chromosom sind weiblich, alle Tiere mit mindestens einem Y-Chromosom sind männlich. Also sind im Normalfall XX-Katzen wie in Ab-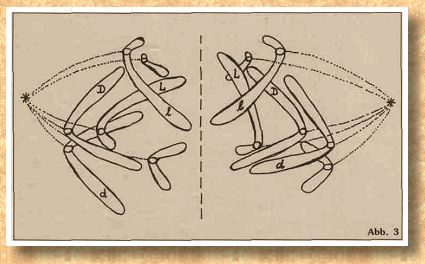 Neu sind die Pfeile, die Ihnen lediglich eine Vorstellung davon geben sollen, wie die Spindelfasern ansetzen und in welche Richtung diese ziehen werden. Wir nehmen wieder vereinfacht an, daß an jedem Zentromer nur zwei Spindelfasern ansetzen, deren Zugrichtung zu genau entgegengesetzten Zellpolen weist. Beim natürlichen Vorgang sind es ganze Bündel von Spindelfaser, die an den Zentromeren zerren. Wenn dann die Zentromere geteilt sind und die beiden Spalthälften oder Chromatiden eines jeden Chromosoms auseinanderweichen, dann bilden sich auch dazwischen noch Spindelfasern, die die beiden Chromatiden auseinanderdrücken. Dann gibt es auch noch Fasern, die direkt von Zellpol zu Zellpol verlaufen und sozusagen „Schienen" bilden, auf denen die Chromatiden zu den Polen „gelenkt" werden. Also bleiben wir lieber bei unserem einfachen Schema und kommen wir mit Abbildung 3 zur nächsten Phase der Mitose, der Anaphase. In der Anaphase werden die beiden Chromatiden eines jeden Chromosoms mit Hilfe der Spindelfasern voneinander getrennt und wandern zu gegenüberliegenden Zellpolen. Sobald die Chromatiden an den Zellpolen angekommen sind, wird eine neue Zellwand gebildet (gestrichelte Linie). Damit sind zwei Tochterzellen entstanden, die, wie Sie leicht selber nachprüfen können, genau die gleiche Gen- und Allelenausstattung haben, wie die Ausgangszelle. Die Chromosomen sind lediglich halb so dick, weil sie nur noch jeweils aus einer Chromatide bestehen. Es folgt die Telephase: Die Chromosomen beginnen sich zu entspiralisieren und in beiden Tochterzellen wird eine neue Kernmembran aufgebaut.
Neu sind die Pfeile, die Ihnen lediglich eine Vorstellung davon geben sollen, wie die Spindelfasern ansetzen und in welche Richtung diese ziehen werden. Wir nehmen wieder vereinfacht an, daß an jedem Zentromer nur zwei Spindelfasern ansetzen, deren Zugrichtung zu genau entgegengesetzten Zellpolen weist. Beim natürlichen Vorgang sind es ganze Bündel von Spindelfaser, die an den Zentromeren zerren. Wenn dann die Zentromere geteilt sind und die beiden Spalthälften oder Chromatiden eines jeden Chromosoms auseinanderweichen, dann bilden sich auch dazwischen noch Spindelfasern, die die beiden Chromatiden auseinanderdrücken. Dann gibt es auch noch Fasern, die direkt von Zellpol zu Zellpol verlaufen und sozusagen „Schienen" bilden, auf denen die Chromatiden zu den Polen „gelenkt" werden. Also bleiben wir lieber bei unserem einfachen Schema und kommen wir mit Abbildung 3 zur nächsten Phase der Mitose, der Anaphase. In der Anaphase werden die beiden Chromatiden eines jeden Chromosoms mit Hilfe der Spindelfasern voneinander getrennt und wandern zu gegenüberliegenden Zellpolen. Sobald die Chromatiden an den Zellpolen angekommen sind, wird eine neue Zellwand gebildet (gestrichelte Linie). Damit sind zwei Tochterzellen entstanden, die, wie Sie leicht selber nachprüfen können, genau die gleiche Gen- und Allelenausstattung haben, wie die Ausgangszelle. Die Chromosomen sind lediglich halb so dick, weil sie nur noch jeweils aus einer Chromatide bestehen. Es folgt die Telephase: Die Chromosomen beginnen sich zu entspiralisieren und in beiden Tochterzellen wird eine neue Kernmembran aufgebaut.